
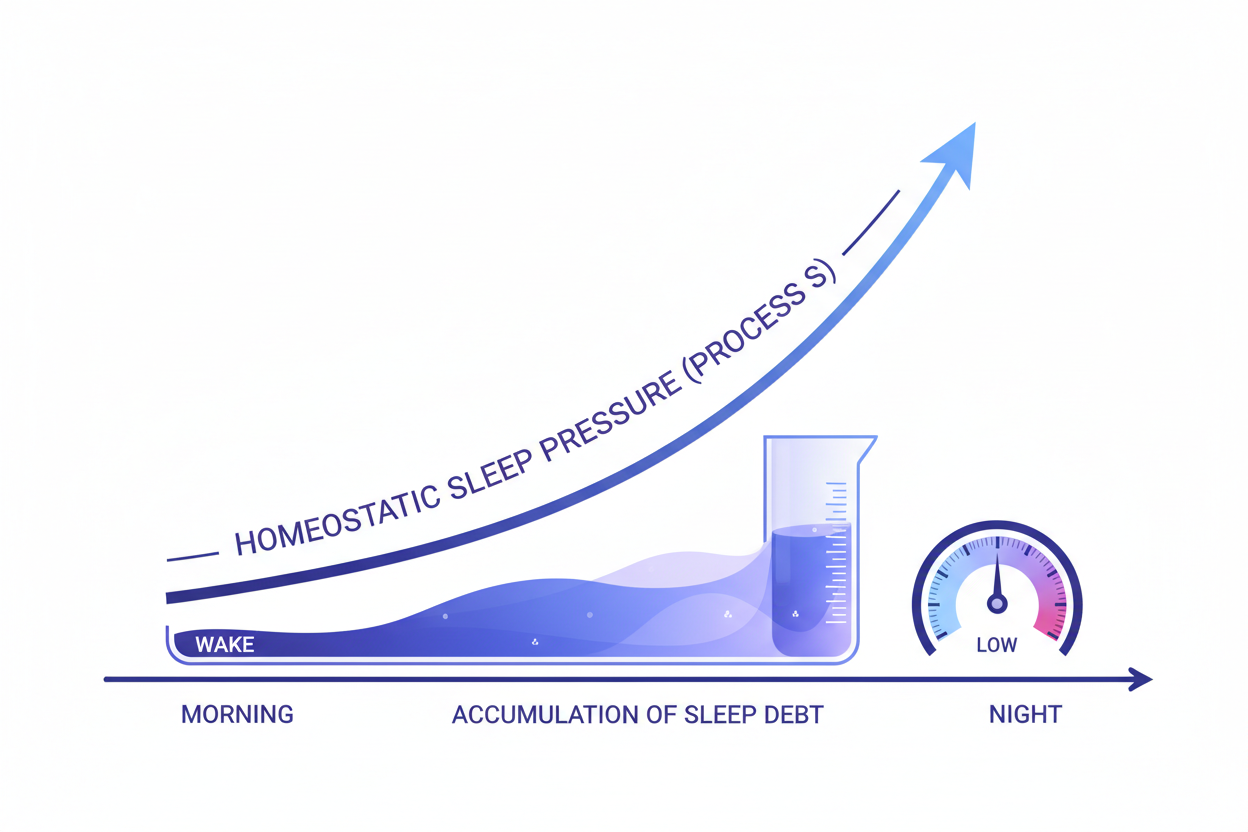
In 1982, Swiss sleep researcher Alexander Borbély published a paper in Human Neurobiology that would become the most influential theoretical framework in sleep science: a mathematical two-process model proposing that human sleep timing and duration are governed not by a single biological mechanism but by the interaction of two entirely independent regulatory systems. Process S — the homeostatic sleep drive — accumulates as a rising pressure during every hour of wakefulness, peaks before sleep onset, and dissipates exponentially during sleep. Process C — the circadian timing signal — is an alerting drive generated by the suprachiasmatic nucleus (SCN) of the hypothalamus that oscillates on a near-24-hour cycle regardless of how long the person has been awake, peaking in the early evening hours and falling through the night. Sleep occurs, Borbély's model predicted, when Process S rises above Process C — when homeostatic pressure overcomes the circadian alerting signal. The elegance of this model was that it made specific, testable predictions about every common sleep disturbance that clinical observation had documented: why shift workers cannot sleep deeply at the wrong circadian phase even when severely sleep-deprived (Process S high, Process C also high — the two processes do not converge); why a brief afternoon nap sharply reduces evening sleepiness (Process S partially dissipated); why sleep becomes lighter and more fragmented in midlife (the amplitude of both Process S and Process C diminishes with age).
This Understanding StAR protein establishes the first principle of mechanistically coherent men's vitality supplementation: supporting testosterone production is not fundamentally about providing testosterone precursors or stimulating LH signalling. It is about maintaining the mitochondrial capacity that Leydig cell steroidogenesis requires at its molecular foundation. This reframing opens a clinical picture of male hormonal health that is considerably more nuanced than total serum testosterone measurements alone reveal — one that includes the SHBG dynamics that determine how much of any testosterone produced is actually bioavailable to tissues, the sperm mitochondrial architecture whose function determines fertility parameters independently of hormonal status, and the oxidative mechanisms that impair both steroidogenesis and sperm DNA integrity in ways that specific antioxidant nutrients address with documented, measurable efficacy. This article covers five dimensions of male vitality biology that have not been addressed as primary subjects in the Zenutri article series, mapping each to the TGA-listed formulations whose mechanisms are directly relevant.because it defines what a nutritional sleep intervention actually needs to accomplish — and reveals immediately why no single supplement can be universally effective regardless of why a person is struggling with sleep. Someone whose Process S accumulation is impaired (perhaps because they are sedentary, sleep-deprived from the night before, or consuming adenosine-blocking caffeine through the afternoon) needs different support from someone whose Process C circadian signal is dysregulated (delayed chronotype from evening screen use, disrupted NAD+/SIRT1 circadian clock fidelity from age, or impaired melatonin onset). This article provides the Two-Process Model as the clinical framework, then covers the molecular mechanisms of each process in detail — the adenosine homeostatic chemistry of Process S, the SCN transcription-translation feedback loop of Process C, and the glycine thermoregulatory mechanism and melatonin receptor pharmacology that bridge them — mapping each to the specific Zenutri TGA-listed formulations that the evidence identifies as relevant.
Key Takeaways
- Understand Borbély's Two-Process Model of sleep regulation — the interaction between Process S (homeostatic sleep pressure, driven by progressive adenosine accumulation during wakefulness in basal forebrain circuits as a metabolic by-product of neuronal ATP activity) and Process C (the SCN circadian alerting signal generated by the CLOCK/BMAL1/CRY/PER transcription-translation feedback oscillator) — and why identifying which process is disrupted in your specific sleep presentation determines which nutritional support will be mechanistically relevant rather than merely sedating.
- Discover the adenosine homeostatic sleep pressure chemistry — how adenosine accumulates extracellularly in the basal forebrain and anterior hypothalamus during wakefulness by dephosphorylation of AMP to adenosine via ecto-5'-nucleotidase (CD73), how adenosine A1 and A2A receptors in sleep-promoting VLPO neurons inhibit the wake-promoting histaminergic tuberomammillary nucleus and cholinergic basal forebrain, and why the magnesium-dependent ATP synthesis rate determines both the pace of adenosine accumulation and the efficiency of its nocturnal clearance through the glymphatic system.
- Learn the SCN molecular clock: CLOCK/BMAL1 transcription-translation feedback loop — the heterodimer of CLOCK and BMAL1 that drives transcription of Period (PER1, PER2, PER3) and Cryptochrome (CRY1, CRY2) genes, the progressive accumulation of PER/CRY repressor complexes that inhibit their own transcription after a time delay generating the ~24-hour oscillation, and why NAD+/SIRT1 enzymatic activity is required to maintain the post-translational modification precision of this clock — making nicotinamide riboside (NR) a mechanistically coherent circadian support supplement rather than simply a longevity nutrient.
- Understand glycine thermoregulatory sleep induction — the Bannai and Makino 2012 clinical evidence from Sleep and Biological Rhythms demonstrating that oral glycine before sleep improves sleep quality, reduces fatigue on waking, and enhances daytime performance through NMDA receptor partial agonism in peripheral vascular smooth muscle that produces core body temperature reduction — the prerequisite for slow-wave sleep entry — and why magnesium glycinate (MagLipo Core AUST L 520793) delivers this mechanism alongside its direct NMDA/GABA sleep architecture support.
- Apply the MT1/MT2 melatonin receptor pharmacology and dim light melatonin onset (DLMO) timing framework — the differential roles of MT1 (acute sleep-promoting inhibition of SCN firing) and MT2 (circadian phase-shifting) receptors, the importance of DLMO as the true circadian phase marker that should anchor supplement timing rather than clock time, why Vitamin D3 morning supplementation supports the daytime serotonin → melatonin precursor pathway while evening D3 may delay DLMO in sensitive individuals, and the complete Zenutri Metabolic Cofactor Bundle (Reversa NR AUST L 520794 + MagLipo Core AUST L 520793, $50 AUD) as the two-mechanism Process S/Process C sleep protocol.
The Two-Process Model: Process S, Adenosine, and the Homeostatic Sleep Drive
Process S — the homeostatic sleep pressure component of Borbély's model — is the sleep drive you feel as a growing urgency to sleep the longer you remain awake. It is not a feeling of tiredness in any vague sense; it has a specific, well-characterised neurochemical substrate. The primary molecular mediator of Process S is adenosine, an inhibitory purine nucleoside produced as a metabolic by-product of neuronal ATP activity during wakefulness. Every action potential, every synaptic event, every metabolic process in a waking neuron that consumes ATP produces ADP and AMP, which ecto-5'-nucleotidase (the CD73 enzyme expressed on the extracellular surface of neurons and astrocytes) dephosphorylates to adenosine. Adenosine accumulates extracellularly in the basal forebrain and anterior hypothalamus across the waking day, with concentrations rising in proportion to the duration and intensity of wakefulness — producing the progressive increase in subjective sleepiness and the objective electroencephalographic slow-wave activity that characterise the rising Process S signal.
Adenosine's sleep-promoting action operates primarily through two receptor subtypes. A1 adenosine receptors in wake-promoting cholinergic basal forebrain neurons directly inhibit these cells when adenosine binds, reducing the acetylcholine release that sustains cortical arousal and attentional function. A2A adenosine receptors in the ventral striatum and nucleus accumbens activate sleep-promoting neurons in the ventrolateral preoptic area (VLPO), which in turn inhibit the histaminergic tuberomammillary nucleus and the noradrenergic locus coeruleus — the two primary ascending arousal systems that maintain wakefulness. The accumulating adenosine therefore progressively shifts the balance of activity away from wake-promoting circuits and toward sleep-promoting circuits — driving the convergence of Process S with Process C at sleep onset. Caffeine's mechanism as a sleep-suppressing compound is competitive blockade of both A1 and A2A receptors — effectively preventing adenosine from exerting its sleep-promoting signal without reducing adenosine accumulation, which is why caffeine's wakefulness effect is temporary and followed by a rebound of intense sleepiness when it wears off and the accumulated adenosine can finally bind.
The Glymphatic System: Adenosine Clearance During Sleep
The clearance of accumulated adenosine during sleep is not simply a passive dissipation process — it occurs partly through the glymphatic system, a brain-specific waste clearance architecture identified by Maiken Nedergaard's group at the University of Rochester in 2013. During non-REM slow-wave sleep, aquaporin-4 water channels on astrocytic endfeet (the specialised projections that wrap around brain blood vessels) drive a convective flow of cerebrospinal fluid through the interstitial space of the brain that clears metabolic waste products — including adenosine and its metabolic products — at substantially higher rates than during wakefulness. The glymphatic system's activity is highest during the deepest stage of slow-wave sleep (N3/delta sleep), creating a positive reinforcement loop: adequate adenosine accumulation drives deeper slow-wave sleep, which activates more efficient glymphatic adenosine clearance, which restores the waking brain's adenosine baseline for the following day's Process S pressure. Anything that reduces the depth or duration of slow-wave sleep — including magnesium insufficiency, alcohol (which suppresses slow-wave sleep despite initial sedation), or elevated cortisol at sleep onset — impairs both the restoration of cognitive function and the efficiency of adenosine clearance, producing the compound fatigue of consecutive nights of poor sleep quality. Taking the free Zenutri health quiz can help determine whether sleep quality is linked to broader nutritional deficiencies in your profile.
The SCN Molecular Clock: CLOCK/BMAL1 and the Circadian Oscillator
The suprachiasmatic nucleus (SCN) — a pair of bilateral nuclei sitting just above the optic chiasm in the hypothalamus, containing approximately 20,000 neurons in total — is the master circadian pacemaker whose output constitutes Process C in Borbély's model. The SCN generates a self-sustaining oscillation with a near-24-hour period through a molecular mechanism: the transcription-translation feedback loop (TTFL) that is among the most evolutionarily conserved biochemical clocks in biology, present in organisms from cyanobacteria to humans.
The core TTFL operates as follows. The CLOCK protein heterodimerises with BMAL1 (Brain and Muscle ARNT-Like protein 1) in the cytoplasm; the CLOCK/BMAL1 heterodimer translocates to the nucleus and binds E-box promoter elements (CACGTG consensus sequences) in the promoter regions of the Period (PER1, PER2, PER3) and Cryptochrome (CRY1, CRY2) genes, driving their transcription. PER and CRY proteins are synthesised in the cytoplasm, form PER/CRY repressor complexes, and translocate back into the nucleus — where they inhibit the CLOCK/BMAL1 complex, suppressing their own transcription. This self-inhibition creates the time delay required for oscillation: as PER/CRY concentrations rise, transcriptional drive falls; as PER/CRY are progressively phosphorylated by CK1ε and CK1δ kinases and targeted for proteasomal degradation, transcriptional inhibition is relieved and the cycle begins again. The full period of this feedback loop — transcription, translation, nuclear translocation, inhibition, degradation, and relief — takes approximately 24 hours under normal conditions.
The precision of this molecular oscillator depends critically on the post-translational modification steps — the phosphorylation of PER proteins by CK1 kinases, the acetylation of BMAL1 by CLOCK's acetyltransferase activity, and the deacetylation of both PER2 and BMAL1 by SIRT1. SIRT1 is the NAD+-dependent deacetylase whose activity is entirely dependent on adequate cellular NAD+ — making the NAD+ pool a direct determinant of molecular clock fidelity. When NAD+ is depleted (as occurs progressively with age through the PARP-1 and CD38 drain mechanisms covered in the anti-ageing article), SIRT1 activity falls, BMAL1 acetylation accumulates, and the CLOCK/BMAL1 transcriptional drive is impaired — producing the gradual loss of circadian rhythm amplitude, reduced melatonin onset precision, and fragmented sleep architecture that many Australian adults in midlife and beyond experience as progressive sleep quality deterioration. This is the mechanistic basis for the circadian-restorative rationale for nicotinamide riboside (NR) supplementation that extends beyond its longevity applications — maintaining the NAD+/SIRT1 axis that molecular clock precision depends on.
Light Entrainment Through the Retinohypothalamic Tract
The SCN's near-24-hour oscillation must be entrained to exactly 24 hours each day by environmental light input. This entrainment occurs through the retinohypothalamic tract (RHT) — a monosynaptic neural projection from intrinsically photosensitive retinal ganglion cells (ipRGCs) expressing the photopigment melanopsin (peak absorption 480nm, blue-wavelength light) directly to the SCN. Morning light exposure activates ipRGCs, which release glutamate and PACAP (pituitary adenylate cyclase-activating polypeptide) at the SCN, resetting the molecular clock to align with the external 24-hour day. Evening blue-wavelength light exposure from screens and LED lighting delays this entrainment signal — the practical basis for recommendations to reduce screen use in the 1-2 hours before sleep. For Australian adults whose sleep timing is delayed (evening chronotype or social jet lag), supporting both the internal molecular clock precision through NAD+/SIRT1 and the external entrainment quality through morning light exposure and evening light reduction addresses the root Process C dysregulation rather than simply masking symptoms with sedating supplements.
Glycine Thermoregulation, Core Body Temperature, and Slow-Wave Sleep Entry
The requirement for a fall in core body temperature (CBT) at sleep onset is one of the most reliable and mechanistically well-understood aspects of human sleep physiology. Core body temperature must decline by approximately 1-2°C from its late-afternoon peak to its nocturnal nadir to facilitate the transition from wakefulness into slow-wave sleep — a thermoregulatory event driven by peripheral vasodilation in the skin and extremities (hands and feet) that transfers heat from the body core to the environment. Impaired peripheral heat dissipation — from cold environments, Raynaud's phenomenon, or sympathetic nervous system overactivation, maintaining peripheral vasoconstriction — delays sleep onset and reduces slow-wave sleep depth, even when adenosine accumulation (Process S) is adequate. This thermoregulatory requirement is the mechanistic basis for the clinical finding that warm baths or foot warming 1-2 hours before sleep improve sleep onset latency — both work by accelerating peripheral vasodilation and heat dissipation, lowering CBT toward the sleep-onset threshold.
Glycine exerts a sleep-promoting effect through precisely this thermoregulatory mechanism. The 2012 clinical trial by Bannai and Makino, published in Sleep and Biological Rhythms, administered 3g oral glycine to healthy volunteers 1 hour before bedtime and measured both objective polysomnographic and subjective sleep outcomes. Participants receiving glycine showed significant reductions in sleep onset latency, higher subjective sleep quality scores, reduced fatigue on waking assessed by the Fatigue Assessment Scale, and improved daytime cognitive performance on memory recognition tasks. The mechanism identified through core body temperature monitoring: glycine-activated NMDA receptors at the glycine co-agonist site in peripheral vascular smooth muscle and spinal cord interneurons, producing peripheral vasodilation — measurably increasing peripheral skin blood flow and accelerating CBT reduction toward the sleep-onset threshold. The effect was physiological rather than pharmacological: glycine was facilitating the body's own thermoregulatory sleep mechanism rather than suppressing CNS function from outside.
What makes this glycine finding particularly relevant for the Zenutri protocol is that MagLipo Core (AUST L 520793) provides magnesium as an amino acid chelate — specifically magnesium glycinate, in which magnesium is bound to glycine through a coordinate covalent bond. The glycine released from this chelate during intestinal absorption and metabolic processing is physiologically available and meaningful: while 3g pure glycine was used in the Bannai trial, the glycine component of magnesium glycinate contributes incrementally to glycine availability alongside any dietary glycine intake. More importantly, the magnesium from the chelate provides its own independent sleep-supporting mechanism through NMDA receptor antagonism and GABA-A potentiation — so MagLipo Core's sleep benefit operates through two converging mechanisms (magnesium NMDA/GABA and glycine thermoregulation) through a single formulation. This mechanistic layering distinguishes it from standalone magnesium oxide or inorganic zinc supplements that lack a glycine component. For Australian adults experiencing difficulty with sleep onset specifically — the most common presentation of the thermoregulatory mechanism's impairment — MagLipo Core taken 30-60 minutes before intended sleep provides the most directly mechanism-aligned support in the current Zenutri TGA-listed range.
MT1/MT2 Receptor Pharmacology, DLMO, and the Melatonin Timing Framework
Melatonin is the pineal gland's molecular signal of biological darkness — synthesised from tryptophan through a two-step enzymatic pathway (tryptophan → serotonin by tryptophan hydroxylase and aromatic amino acid decarboxylase; serotonin → N-acetylserotonin → melatonin by arylalkylamine N-acetyltransferase, AANAT, and hydroxyindole-O-methyltransferase, HIOMT) whose rate-limiting enzyme AANAT is activated by SCN-driven noradrenergic innervation of the pineal in the absence of light. Melatonin secretion begins when the SCN's light-suppression signal diminishes at dusk, typically rising 2-3 hours before habitual sleep onset — the timing point called dim light melatonin onset (DLMO) that represents the most accurate, non-invasive marker of the individual's current circadian phase. DLMO is typically measured in research settings from salivary melatonin samples taken under dim light conditions (less than 10 lux) across the evening, but its existence as a precisely timed biological event is relevant to anyone planning sleep-supporting supplementation: it defines when the body's internal sleep biology is activating, and aligning nutritional interventions with this timing window maximises their mechanistic synergy with endogenous sleep regulatory biology.
Melatonin's physiological effects are mediated through two G-protein-coupled receptor subtypes that have distinct functional roles. The MT1 receptor (encoded by MTNR1A) is expressed in the SCN and pars tuberalis of the hypothalamus; its activation by melatonin produces acute inhibition of SCN neuronal firing rate — the mechanism by which endogenous melatonin onset signals the beginning of biological night to the master clock and directly promotes sleep onset. MT2 receptors (MTNR1B) are also expressed in the SCN mediate phase-shifting effects: melatonin acting at MT2 receptors shifts the phase of the CLOCK/BMAL1 oscillator in a direction-dependent manner based on the timing of the signal relative to the endogenous DLMO — early melatonin (before DLMO) advances the clock; late melatonin (after usual sleep time) delays it. This phase-shifting pharmacology through MT2 is the basis for chronobiotic applications of melatonin supplementation — using low-dose melatonin timed to advance or delay the circadian phase rather than simply to sedate. For Australian adults with delayed chronotype (DLMO occurring late, after 11pm), the combination of morning Vitamin D3 (supporting daytime serotonin synthesis, the melatonin precursor pool for the coming night) and NR-supported NAD+/SIRT1 circadian clock fidelity addresses the molecular causes of late DLMO rather than simply accepting the delayed phase as fixed.
Vitamin D3 and the Serotonin-Melatonin Precursor Pathway
Vitamin D3 (cholecalciferol) contributes to sleep architecture through the melatonin precursor pathway in a way that is distinct from its better-known immune and bone mineralisation functions. Serotonin — the neurotransmitter synthesised from tryptophan by tryptophan hydroxylase (TPH) in brainstem raphe neurons and by TPH1 in the gut — is the immediate precursor of melatonin in the pineal gland. The TPH enzyme's activity is regulated in part by calcitriol (active Vitamin D): the promoter region of the TPH2 gene contains a functional vitamin D response element (VDRE), and adequate Vitamin D3 status supports the daytime serotonin synthesis that determines the melatonin precursor pool available to the pineal gland in the evening. Australian adults who are Vitamin D insufficient (31% of the population, as established in the vitamin D deficiency article) may have subtly impaired serotonin synthesis, which reduces melatonin precursor availability — one mechanism linking the widespread Australian Vitamin D insufficiency to the sleep quality complaints that are similarly prevalent in population surveys. Osteo+Core (AUST L 520792) provides D3 at 1,000 IU taken in the morning — the timing that supports daytime serotonin synthesis for the coming evening's melatonin onset while avoiding any potential evening DLMO delay from late D3 dosing. The K2 MK-7 at 180mcg in the same formulation has emerging sleep-quality data independent of the bone and cardiovascular evidence that remains its primary clinical evidence base.
The Zenutri Sleep Protocol: Process S and Process C Support
The Two-Process Model provides the architecture for a mechanistically coherent sleep-supplement protocol: address the homeostatic sleep drive (Process S) with magnesium amino acid chelate plus glycine for thermoregulation in the evening; support the circadian clock precision (Process C) with NAD+/SIRT1 and Vitamin D3/serotonin pathway supplementation in the morning. These two timing windows map to the two distinct Zenutri formulations whose mechanisms are most directly relevant.
Evening (30-60 minutes before intended sleep, after reducing light exposure):
MagLipo Core (AUST L 520793) — Magnesium amino acid chelate 55mg elemental, Alpha-lipoic acid 150mg.
Addresses sleep through: Magnesium NMDA receptor antagonism reducing the glutamatergic cortical arousal drive that prevents slow-wave sleep transition; GABA-A positive modulation supporting inhibitory GABAergic sleep-onset tone; glycine thermoregulatory peripheral vasodilation lowering core body temperature for slow-wave sleep entry (Bannai 2012); ALA antioxidant protection supporting mitochondrial ATP synthesis efficiency that determines both the pace of adenosine accumulation (Process S pressure) and glymphatic clearance quality during deep sleep. Note: ALA has broad-spectrum antioxidant activity; individuals on antidiabetic medications (ALA influences insulin sensitivity) or thyroid medications should confirm with their GP before initiating. Take with a small amount of food to optimise absorption of magnesium amino acid chelate.
Morning (with breakfast, alongside light exposure if possible):
Reversa NR (AUST L 520794) — Nicotinamide Riboside 150mg, Resveratrol 75mg, Magnesium amino acid chelate 55mg, BioPerine 6.95mg.
Addresses sleep through: NAD+ restoration via the NRK pathway (Brenner 2018, Nature Communications — 40-90% blood NAD+ elevation at 100-300mg NR) supporting SIRT1 enzymatic activity for CLOCK/BMAL1 post-translational modification precision; SIRT1-mediated deacetylation of PER2 and BMAL1 maintaining molecular clock fidelity; resveratrol SIRT1 allosteric activation maximising the circadian utility of restored NAD+. Safety: piperine CYP3A4 interaction for antidepressant medication users (consult GP); pregnancy — pause (see maternity article); warfarin users — warfarin interaction monitoring advised.
Osteo+Core (AUST L 520792) — D3 1,000 IU, K2-MK7 180mcg.
Addresses sleep through: D3 VDR-VDRE regulation of TPH2 gene expression supporting daytime serotonin synthesis for the melatonin precursor pool; D3 general circadian entrainment support. Morning timing critical — supports daytime serotonin synthesis and avoids potential evening DLMO delay. K2 warfarin interaction — GP discussion warranted for anticoagulant users.
The core two-formulation sleep protocol — Zenutri Metabolic Cofactor Bundle (Reversa NR AUST L 520794 + MagLipo Core AUST L 520793, $50 AUD, save 24%) — addresses both Process S homeostatic sleep pressure (evening magnesium/glycine) and Process C circadian clock precision (morning NR/SIRT1) in the most streamlined two-supplement sleep support protocol available in the Zenutri TGA-listed range. Adding Osteo+Core for the D3 melatonin precursor pathway completes the three-formulation protocol for adults with confirmed Vitamin D insufficiency or prominent sleep-onset delay that may reflect impaired melatonin onset timing.
For the complete sleep architecture picture — including cortisol rhythm dysregulation and the HPA axis evening cortisol patterns that prevent the natural Process S/Process C convergence at sleep onset — the ashwagandha benefits article covers the withanolide GABAergic and glucocorticoid receptor modulation mechanisms that complement the magnesium/NR protocol for adults whose sleep disruption is primarily driven by elevated evening cortisol. The complete Zenutri Core Nutrient System brings all four formulations together for a comprehensive foundation. Take the free Zenutri health quiz to identify whether your sleep presentation maps primarily to Process S, Process C, or HPA axis dysregulation — and receive a personalised supplement recommendation tailored to your specific sleep architecture.
Sleep Is a Biological Process, Not a Pharmacological Problem
The Two-Process Model, the adenosine homeostatic chemistry of Process S, the SCN CLOCK/BMAL1 molecular clock of Process C, the glycine thermoregulatory mechanism of Bannai 2012, and the MT1/MT2 receptor pharmacology of melatonin timing — together these mechanisms establish that restorative sleep is not a state the body fails to produce without pharmacological intervention. It is a precisely regulated biological programme that the body initiates when the right conditions are present: adenosine accumulation above the sleep threshold, circadian phase alignment between the internal clock and the external environment, core body temperature declining through peripheral vasodilation, and the melatonin onset signal confirming biological night. Nutritional sleep supplements that work do so by supporting the conditions the body requires for this programme to run — not by overriding the nervous system from the outside. Magnesium amino acid chelate and glycine support thermoregulation and GABA/NMDA balance. NR supports the molecular clock fidelity. D3 supports the melatonin precursor pathway. Each works with the biology rather than against it — and each is available in TGA-listed, Australian-made form in the Zenutri range.
Always read the label. Follow the directions for use. Supplements are not a substitute for a balanced diet. If symptoms persist, consult your healthcare professional. This article is for educational purposes and does not constitute individual medical advice.
Explore the Zenutri Metabolic Cofactor Bundle — the Process S and Process C sleep support protocol in one $50 AUD bundle — or take the free health quiz to identify the specific sleep regulatory system most relevant to your presentation and receive a personalised recommendation.
Frequently Asked Questions
What are the best sleep supplements in Australia?
The strongest evidence maps to magnesium amino acid chelate (MagLipo Core AUST L 520793) for slow-wave sleep via NMDA antagonism, GABA-A potentiation, and glycine thermoregulatory core body temperature reduction (Bannai 2012, Sleep and Biological Rhythms); and nicotinamide riboside 150mg (Reversa NR AUST L 520794) for NAD+/SIRT1 circadian clock fidelity and CLOCK/BMAL1 molecular oscillator precision (Brenner 2018). These are combined in the Zenutri Metabolic Cofactor Bundle ($50). Osteo+Core D3 (AUST L 520792) adds the serotonin→melatonin precursor pathway support. All should be confirmed with your GP if you are on medications or managing a diagnosed sleep disorder.
What is the Two-Process Model of sleep?
Borbély's 1982 Two-Process Model identifies two independent systems governing human sleep. Process S (homeostatic sleep pressure) accumulates during wakefulness through progressive adenosine build-up as a neuronal ATP metabolic by-product, rising until it overcomes Process C. Process C is the SCN circadian alerting signal — a CLOCK/BMAL1 transcription-translation feedback oscillator generating a near-24-hour alertness rhythm independent of wakefulness duration. Sleep occurs when Process S exceeds Process C. Most Australian sleep complaints involve dysregulation of one or both processes, and effective sleep supplementation targets the specific process that is impaired rather than broadly sedating.
How does magnesium help with sleep?
Three independent mechanisms: NMDA receptor antagonism, reducing the glutamatergic cortical arousal that prevents slow-wave sleep transition; GABA-A receptor positive modulation, supporting inhibitory sleep-onset tone without receptor dependency; and glycine thermoregulatory peripheral vasodilation (from the glycine amino acid component of magnesium glycinate) lowering core body temperature to the sleep-onset threshold, confirmed by Bannai 2012 in humans. The clinical evidence was generated with highly bioavailable amino acid chelate forms — the 4% absorption rate of magnesium oxide cannot achieve the intracellular concentrations these mechanisms require, making form selection the most important quality variable for magnesium sleep supplements. MagLipo Core (AUST L 520793) provides magnesium amino acid chelate at 55mg elemental magnesium.
What does glycine do for sleep?
Glycine exerts sleep-promoting effects through peripheral thermoregulation rather than direct CNS sedation. Bannai and Makino (2012) confirmed that oral glycine reduces sleep-onset latency, improves subjective sleep quality, reduces waking fatigue, and enhances daytime cognitive performance in humans. The mechanism: glycine acts at NMDA receptor glycine co-agonist sites in peripheral vascular smooth muscle, producing peripheral vasodilation, increasing skin blood flow, and accelerating the core body temperature reduction required for slow-wave sleep entry. As one of the two amino acids in magnesium glycinate (MagLipo Core), glycine contributes to this thermoregulatory mechanism alongside magnesium's NMDA/GABA sleep architecture support.
What is DLMO, and why does it matter for supplement timing?
Dim light melatonin onset (DLMO) is the circadian phase marker at which pineal melatonin secretion begins — typically 2-3 hours before habitual sleep onset under dim-light conditions. It represents the body's transition into biological night as signalled by the CLOCK/BMAL1 oscillator through SCN-mediated noradrenergic activation of the pineal AANAT enzyme. DLMO timing determines when sleep-supporting evening supplements (magnesium, glycine) are most mechanistically relevant. Vitamin D3 should be taken in the morning to support daytime serotonin → melatonin precursor synthesis without potentially delaying DLMO in the evening. Adults with consistently delayed DLMO (late chronotype, after 11pm) may benefit from morning NR supplementation to support NAD+/SIRT1 circadian clock fidelity and improve molecular clock precision.
How does nicotinamide riboside help with sleep?
NR supports sleep through the NAD+/SIRT1 molecular clock precision mechanism. The CLOCK/BMAL1 circadian oscillator requires SIRT1 deacetylase activity — entirely NAD+-dependent — to perform the post-translational modification of PER2 and BMAL1 that maintains the ~24-hour period precision of the transcription-translation feedback loop. As NAD+ declines with age (through the PARP-1/CD38 dual drain detailed in the anti-ageing article), SIRT1 activity falls and molecular clock amplitude degrades — contributing to the fragmented sleep architecture and blunted melatonin onset precision that many midlife Australians experience. Reversa NR (AUST L 520794) provides 150mg NR, confirmed by Brenner 2018 to raise blood NAD+ by 40-90%, with resveratrol allosteric SIRT1 activation maximising circadian clock support from the restored NAD+ pool.
Is it safe to take magnesium every night for sleep?
Magnesium amino acid chelate, at doses providing up to 350mg of elemental magnesium daily, is within the NHMRC 2017 safe upper intake level for supplemental magnesium in adults. Unlike benzodiazepines or Z-drugs, magnesium does not produce receptor tolerance, dependency, or next-day cognitive impairment — its sleep-supporting mechanisms operate through physiological NMDA/GABA modulation that does not require escalating doses or produce withdrawal effects. For adults with renal impairment, high-dose magnesium supplementation should be confirmed with a GP as renal clearance of magnesium is the primary safety consideration. MagLipo Core (AUST L 520793) provides 55mg elemental magnesium per dose — well within safe limits and appropriate for nightly use as part of a sustained sleep support protocol.
Ready to take action on your health?


{kind=link}
اترك تعليقًا
This site is protected by hCaptcha and the hCaptcha Privacy Policy and Terms of Service apply.